AlgaeFeed Oil Drops
Sustainable Marine Lipids
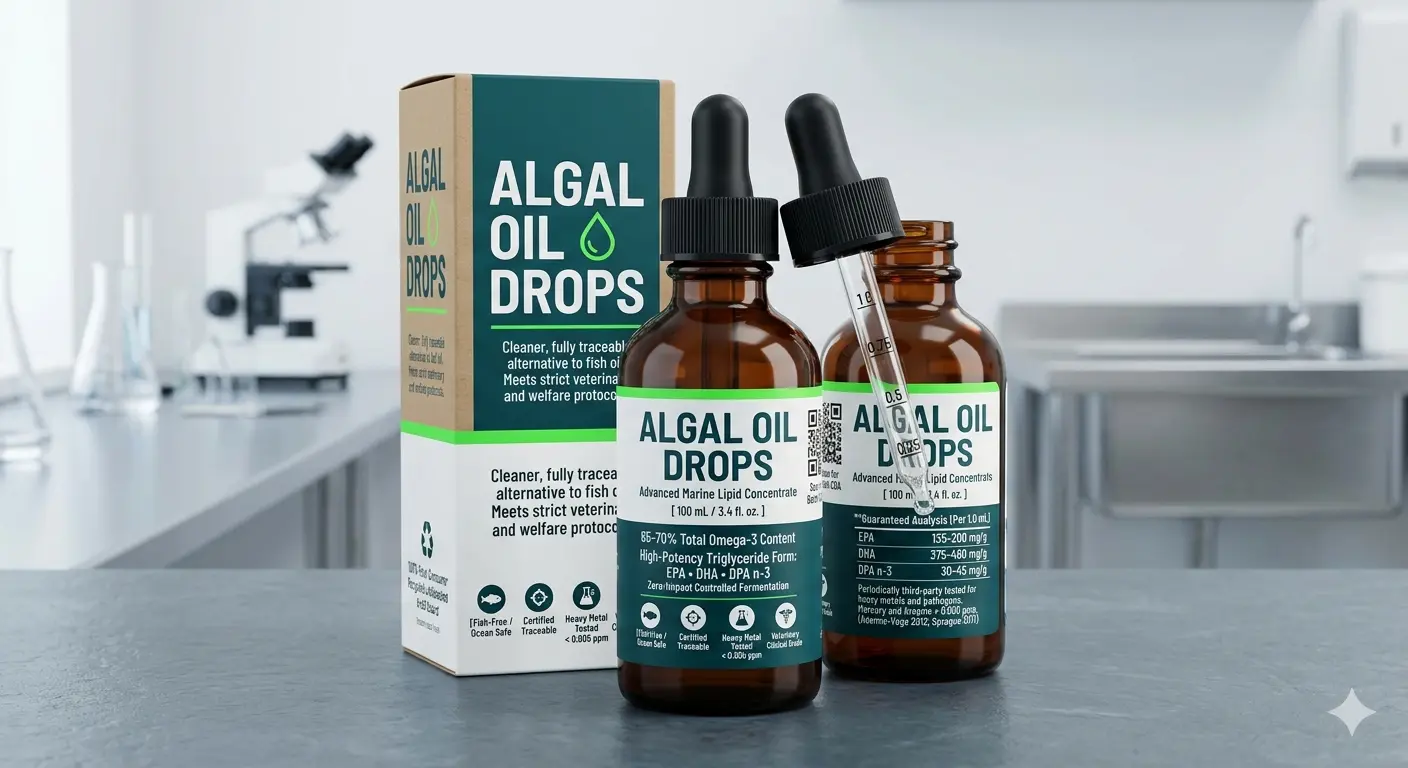
Pure microalgae concentrate: 65–70% total Omega-3. Clean, highly stable nutrition. Zero fish oil.

AlgaeFeed Oil Drops offer a sustainable lipid concentrate delivering highly stable marine nutrition without fish oils or ocean contaminants. Produced via controlled fermentation, it carries none of the oxidative instability or contamination risks associated with traditional fish oils (Adarme-Vega 2012; Sprague 2017).
- Concentrated Omega-3 Profile: Engineered to provide 65–70% total Omega-3 content by peak area in a highly bioavailable format.
- Biologically Active Formats: Rich in preformed long-chain elements including DHA, EPA, and critical DPA n-3.
- Pure Fermentation Matrix: Cultured in bio-secure setups, completely free of industrial ocean pollutants, microplastics, and heavy metals.
- Oxidative Stability: Avoids the rapid breakdown and odor decay that traditionally complicate marine oil logistics.
The Safety & Documentation Standard: For institutional collections and zoological facilities already utilizing standard fish oil supplementation, AlgaeFeed Oil Drops offer a cleaner, fully traceable alternative. Periodically third-party tested for heavy metals and pathogens, with mercury and arsenic remaining below 0.005 ppm alongside non-detectable Salmonella and Listeria parameters, this marine lipid concentrate meets the strict documentation and traceability standards that professional veterinary and welfare protocols demand.
Why Microalgae?
Microalgae is the foundational baseline source of all marine omega-3s. Wild fish and krill do not synthesize these lipids themselves; they merely accumulate them by feeding on algae over time. By sourcing directly from the baseline, AlgaeFeed bypasses the entire aquatic food chain to achieve unmatched purity and nutritional standardizing (Tocher 2015; Sprague 2017).
Traditional marine oils present severe operational challenges. Conventional fish oils suffer from rapid oxidative breakdown, aggressive odor decay, and alarming accumulation risks for environmental heavy metals. Polar krill extraction provides matching active molecules but triggers localized ecosystem disruption alongside heavy commercial supply volatility.
Terrestrial plant substitutes (like flax or chia) are metabolically inadequate for specialized husbandry. They provide omega-3 solely as short-chain ALA. Most birds, reptiles, and exotic managed species lack the metabolic pathway to convert ALA into the required, biologically active long-chain EPA and DHA formats (Klasing 2008; Brenna 2009).
Our premium microalgae matrix is cultured via controlled bio-fermentation. This sterile, inland setup delivers a perfectly consistent fatty acid yield across seasons with zero ocean pollutants, zero microplastics, and structural, protective oxidative stability that fish oils fundamentally lack (Adarme-Vega 2012). This exact biome-isolated process forms the exact foundation of our AlgaeFeed Oil Drops, bottling this raw natural potency into a stable, easy-to-dose lipid architecture.
Why Omega-3? The Core Physiology
Omega-3 fatty acids are indispensable structural components of cell membranes and play a central role in cellular signalling, inflammatory balance, and neurological function across animal systems (Simopoulos 2002; Calder 2017). EPA and DHA represent the biologically active forms required to drive these pathways efficiently.
Critically, many captive species have limited metabolic capacity to convert plant-based short-chain omega-3s, such as alpha-linolenic acid (ALA), into the essential long-chain forms of EPA and DHA (Brenna 2009). This conversion limitation is particularly relevant in avian and reptile nutrition, where conversion efficiency from plant-based precursors is generally low (Klasing 2008).
Without preformed marine lipids added directly to the formulation design, animals kept under managed care often exhibit sub-clinical lipid imbalances that affect tissue resilience, systemic inflammation, and skin barrier maintenance.
| Omega-3 Component | Concentration Range |
|---|---|
| DHA (Docosahexaenoic Acid) | 375 – 460 mg/g |
| EPA (Eicosapentaenoic Acid) | 135 – 200 mg/g |
| DPA n-3 (Docosapentaenoic Acid) | 30 – 45 mg/g |
Synergistic Mineral-Lipid Integration
By pairing the critical mineral density of our AlgaeFeed Mineral Supplement Powder with the targeted soft-tissue support of these oil drops, this dual-action system delivers comprehensive head-to-tail wellness. Together, they form a complete foundation that accelerates skeletal development, ensures healthier sheds and molts, and promotes vibrant plumage and resilient skin (Hand et al.; EFSA 2010–2022).
Whether used for acute professional conditioning or long-term captive care, this synergistic combination allows specialists to deliver precise, concentrated doses that optimize vitality from the inside out (Swanson 2012; NRC 2006).
Help Us Refine the Recipe
We are currently developing this product and would love to hear your feedback. Get in touch to help us refine the final recipe, or to collaborate and take part in the testing phase.

Sources & Scientific References
- Adarme-Vega, T. C., et al. (2012). Microalgae oils as lipid sources for marine organisms and animal nutrition. Progress in Lipid Research, 51(4), 218–226.
- Brenna, J. T., et al. (2009). Alpha-linolenic acid conversion efficiency and its relevance to biological active forms. Prostaglandins, Leukotrienes and Essential Fatty Acids, 80(2-3), 85–91.
- Calder, P. C. (2017). Omega-3 fatty acids and inflammatory processes: From molecules to animals. Proceedings of the Nutrition Society, 76(3), 273–283.
- EFSA NDA Panel (2010–2022). Scientific opinions on algal oil safety parameters as a novel food and feed ingredient matrices. EFSA Journal.
- Hand, M. S., et al. (2010). Small Animal Clinical Nutrition, 5th Edition. Mark Morris Institute.
- Klasing, K. C. (2008). Nutritional energetics and trace lipid conversion paths in avian and reptile nutrition profiles. Journal of Nutrition, 138(2).
- NRC (2006). Nutrient Requirements of Dogs and Cats. National Academies Press. Washington, DC.
- Simopoulos, A. P. (2002). Omega-3 fatty acids in inflammation and chronic disease mitigation frameworks. Journal of the American College of Nutrition, 21(6), 495–505.
- Sprague, M., et al. (2017). Omega-3 fatty acids in aquaculture, microalgae shifts, and broader animal nutrition. Reviews in Fisheries Science & Aquaculture, 25(4), 281–290.
- Swanson, D., et al. (2012). Omega-3 fatty acids and immune function parameters across animal species systems. Advances in Nutrition, 3(1), 1–7.
- Tocher, D. R. (2015). Omega-3 metabolism in fish, primary production source paths, and premium feed ingredients. Fish Physiology and Biochemistry, 41(2), 307–319.